缝隙连接(Gap junctions)又称间隙连接 , 是一种重要的跨细胞通道 , 可以介导分子量小于约1 kD的小分子(如cAMP , IP3 , Glucose等第二信使)或离子(氢离子、钙离子、钾离子等)在细胞间的扩散 。 这种通道广泛地存在于包括脊椎动物和无脊椎动物的各类组织和器官中 , 介导细胞间的化学和电学信号传递 , 从而调控发育、维持稳态 , 保证生物体各个系统的正常工作 。 缝隙连接功能的缺陷与许多疾病有关 , 包括心肌梗死、听力丧失和低髓鞘性疾病等 。
文章图片
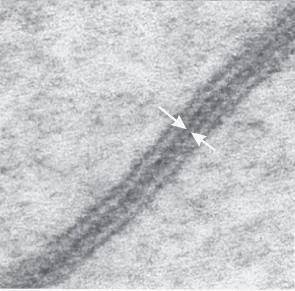
图1. 小鼠肝细胞缝隙连接的电镜观察
(Goodenough, D.A., et al.Cold Spring Harb Perspect Biol, 2009.)
文章图片
(来源:wikipedia)
缝隙连接蛋白
形成缝隙连接的蛋白质在脊椎动物和无脊椎动物中是不同的 。 在脊椎动物中 , 缝隙连接是由连接蛋白(Connexins , Cx)形成的 , 分为5个亚组(α、β、γ、δ或 ε) 。 连接蛋白是根据其预测分子量命名的 , 并根据发现的顺序编号 , 例如 , Cx43大小为~ 43 kd , 是α组的第一个连接蛋白(GJα1) 。 在人类和小鼠基因组中分别已发现21/20种不同的连接蛋白基因 。 在无脊椎动物中 , 缝隙连接蛋白被称为“innexins” 。
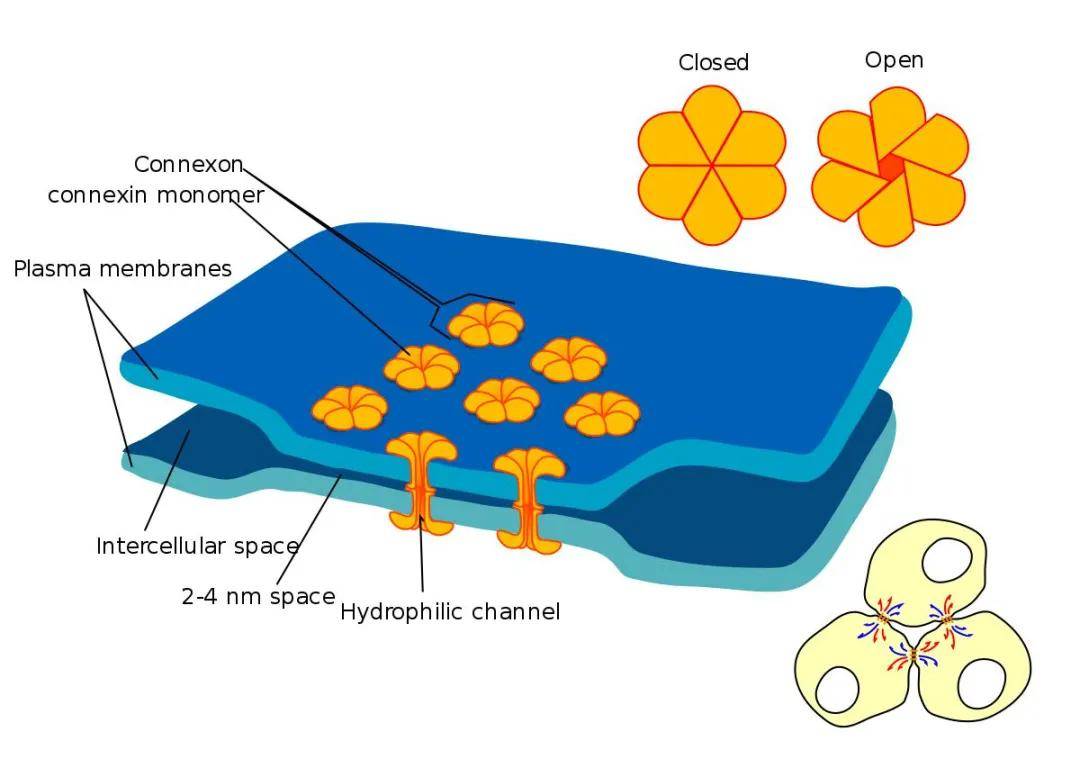
单个连接蛋白在细胞内组装成六聚体 , 称为连接子(connexon)或半通道(hemichannel) 。 由单一连接蛋白组成的连接子称同聚体连接子 , 不同种连接蛋白构成的连接子称异聚体连接子 。 连接子可以与相同连接子对接形成同型缝隙连接 , 也可以与包含不同连接蛋白的连接子对接形成异型缝隙连接 , 从而进一步增加了其组成和功能的多样性 。
缝隙连接位于质膜表面的斑块中 , 由相邻细胞的2个半通道连接而成 , 这些半通道又由6个连接蛋白连接而成 。 连接蛋白具有相似的结构(图3):
?四个跨膜亲水片段TM1、TM2、TM3、TM4 , 为α螺旋结构;
?两个胞外环(Extracellular loop)E1、E2 , 使TM区域TM1-TM2和TM3- TM4具有连续性;
?一个胞质环(cytoplastic loop , CL) , 位于TM2和TM3之间;
?羧基末端(CT)与氨基末端(NT)位于胞质内;
羧基末端差别较大 , 其丝/苏/酪氨酸残基的磷酸化/去磷酸化水平影响缝隙连接的形成及功能状态 。

文章图片
图3. 缝隙连接与单体结构模型
(Joost Willebrords, et al. Cell Commun Adhes, 2016)
连接蛋白存在于兴奋和非兴奋的组织中 , 除了分化的骨骼肌、红细胞和成熟精子细胞之外的所有组织中都有表达 。 连接蛋白编码基因的突变与多种遗传疾病(包括耳聋、神经病、白内障、骨骼异常和皮肤病)的发生相关 , 它们调节免疫反应、细胞增殖、迁移、凋亡和癌变的能力使之成为多种疾病的治疗靶点 。
缝隙连接蛋白的形成和降解
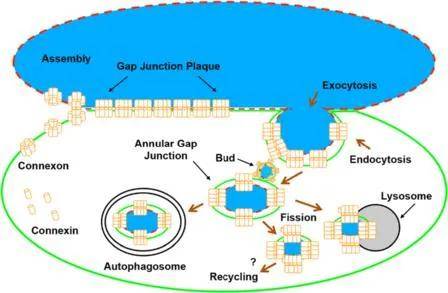
已有研究证明连接蛋白在内质网中合成 , 在高尔基体中加工形成半通道 。 新合成的连接蛋白组装成连接子 , 随后被运输并插入到质膜中 。 在细胞接触过程中 , 连接子与对应细胞的连接子正面对接并聚集形成缝隙连接斑块 。 一旦细胞间通讯不再需要缝隙连接斑块 , 或细胞迁移 , 缝隙连接斑块的一部分(通常是中心部分)或整个斑块内化 , 形成环状缝隙连接 。 环状缝隙连接可以通过一系列过程降解 , 包括溶酶体、自噬体等过程 。 环状缝隙连接囊泡或在环状缝隙连接囊泡降解过程中释放的连接蛋白可能会回到细胞表面循环利用 , 参与新的或现有缝隙连接斑块的形成(图4) 。
文章图片
图4. 缝隙连接斑块的形成及降解示意图
(Falk, M.M., et al. BMC Cell Biol, 2016. )
缝隙连接的研究工具
缝隙连接在各种生理过程中发挥重要作用 , 而在复杂系统(神经系统)的生理或疾病条件下研究缝隙连接耦合需要一种既具有细胞特异性又具有高时空分辨率的非侵入性方法 , 然而以往用于监测缝隙连接的方法包括电生理记录、染料微注射、荧光漂白恢复(FRAP)和分子荧光探针的局部激活(LAMP) , 均缺乏细胞特异性及高时空分辨率 , 并且具有侵入性 , 因此限制了它们的应用 。
为了更好地研究缝隙连接在复杂系统中的分布及生理和病理条件下的功能 , 2019年 , 北京大学李毓龙课题组开发了一种新型的、可基因编码的光遗传学缝隙连接检测探针——PARIS(pairing actuators and receivers to optically isolate gap junctions) , 并将其应用在细胞系、心肌细胞和转基因果蝇的神经系统中重复地检测特定细胞间的缝隙连接通讯 。 该方法首次实现了运用完全遗传编码的方法在特异的细胞类型中非侵入地对缝隙连接通讯进行成像 , 它结合了光学的高度时空操纵性和遗传学的特异性 , 为研究缝隙连接通讯的在体分布、不同生理活动下的功能及调节提供了更多的可能性 。

文章图片
图5. PARIS的作用原理示意图
PARIS包含两部分:第一部分是一个 光控质子“泵”(ArchT) , 它将氢离子输送出细胞;第二部分是一个荧光传感器(质子敏感型荧光蛋白pHluorin , 所用AdV-pHluorinCAAX腺病毒由维真生物提供) , 存在于接收信号的细胞内 , 它对流出的氢离子做出反应 。 将二者分别表达在缝隙连接耦合的细胞中 , 通过光激活质子泵产生跨细胞的质子梯度 , 再通过pHluorin的荧光变化检测两个相邻细胞间的缝隙连接通讯(如图5所示) 。
研究思路
研究人员将探针表达在不同的样品中 , 实验证实PARIS可以在细胞系(HEK293T、HeLa)、新生大鼠心肌细胞以及转基因果蝇的神经系统中可靠、重复地检测缝隙连接通讯 。 基于转基因果蝇的研究结果表明 , PARIS在活体组织中是稳定的 , 并且可以绘制连接特定神经细胞间的缝隙连接 , 并在亚细胞水平揭示了神经元之间通过树突构成的电突触 。
部分研究结果
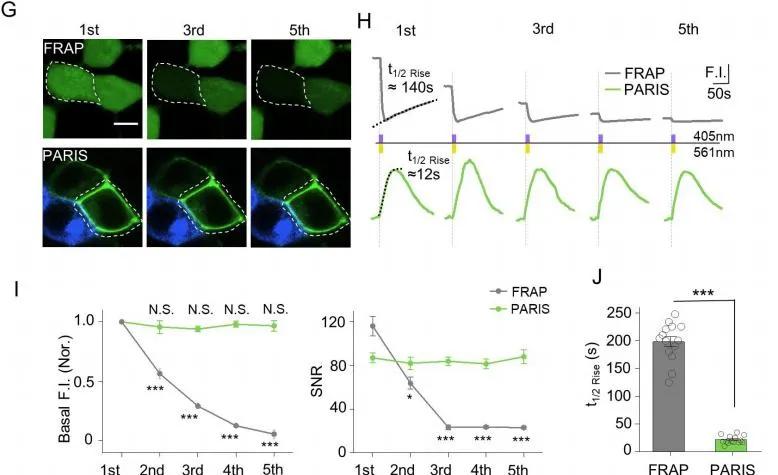
1、PARIS与FRAP(荧光漂白恢复技术)在HEK293T细胞中的功能比较
通过对PARIS与FRAP(基于染料扩散的方法检测缝隙连接介导的光漂白后的荧光恢复)的比较发现 , 相同时间内PARIS信号比FRAP信号更加稳定 , 未曾出现衰减 , 而且PARIS信号的动力学比FRAP快速 , 对应的t1/2分别约为~21s和~197s(图6) 。
文章图片
图6. PARIS与FRAP在HEK293T细胞中的比较
【应用|解锁腺病毒在『缝隙连接』研究中的应用】2、PARIS可检测缝隙连接和由连接蛋白编码基因突变引起的疾病
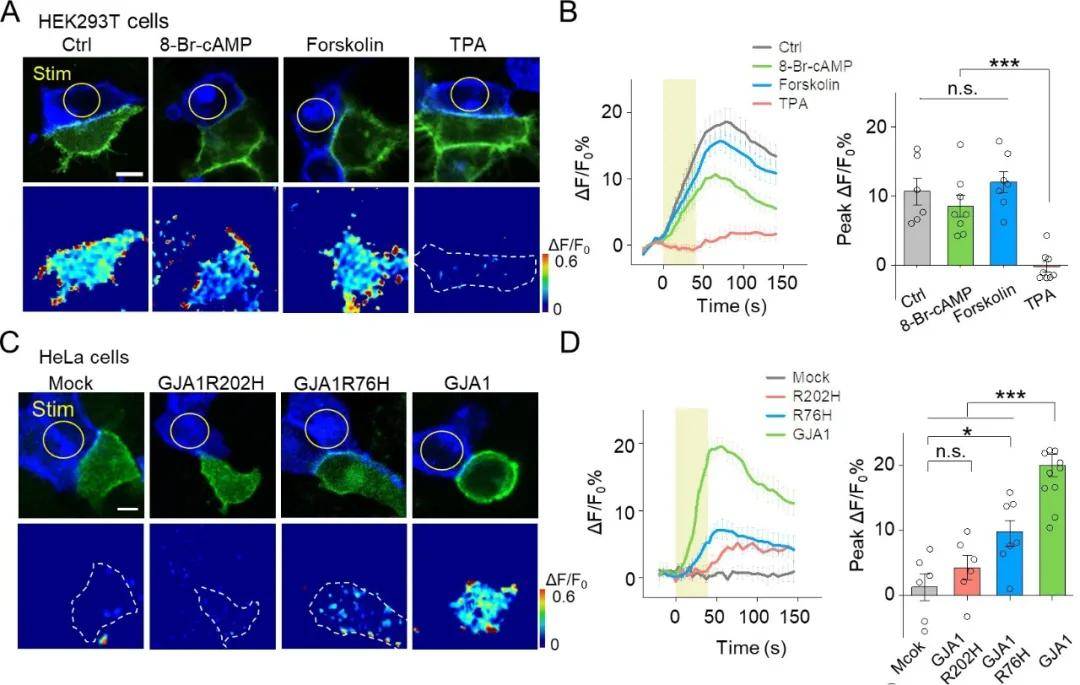
研究表明 , 磷酸化通过影响缝隙连接的组装、降解和门控等途径参与其调节 。 接下来 , 作者探讨了PARIS是否可以在蛋白磷酸化等调节条件下检测缝隙连接:用cAMP类似物8-Br-cAMP、腺苷酸环化酶激动剂Forskolin及蛋白激酶C(PKC)激动剂TPA处理表达PARIS的HEK293T细胞 。 与对照组相比 , 经TPA处理细胞后 , PARIS信号受到显著抑制 , 而8-Br-cAMP和Forskolin的处理对其未造成明显影响 , 表明PKC的激活会抑制缝隙连接 。
已知连接蛋白Cx43编码基因GJA1的突变与许多疾病有关 , 那么PARIS是否可以用来检测连接蛋白编码基因突变引发的疾病呢?作者在不表达内源性连接蛋白的HeLa细胞中表达了PARIS , 光激活“质子泵”细胞时 , 接收器细胞中未产生PARIS信号;而在表达GJA1的HeLa细胞中 , 光激活“质子泵”细胞时 , 相邻接收器细胞中产生了强烈的荧光 。 由于Cx43编码基因GJA1的突变影响了缝隙连接的形成 , 从而使PARIS信号显著降低 。 上述结果表明 , PARIS可用于检测缝隙连接蛋白编码基因突变造成的疾病(图7) 。
文章图片
图7. PARIS可检测缝隙连接和由连接蛋白基因突变引起的疾病
更多精彩内容请关注【维真生物】官网 或 公众号进行查看!
推荐阅读







- 区块|面向2030:影响数据存储产业的十大应用(下):新兴应用
- 下架|APK Installer 和 WSATools 同时躺枪:冒牌应用登陆微软应用商店
- 苏宁|可循环包装规模化应用 苏宁易购绿色物流再上新台阶
- 软件和应用|AcrylicMenus:让Windows 10右键菜单获得半透明效果
- Tencent|原生版微信上架统信UOS应用商店:适配X86、ARM、LoongArch架构
- 技术|使用云原生应用和开源技术的创新攻略
- 飞腾|原生版微信登陆统信UOS应用商店,已适配X86/ARM/LoongArch架构
- 智能化|龙净环保:智能型物料气力输送系统的研究及应用成果通过鉴定
- 软件和应用|iOS/iPadOS端Telegram更新:引入隐藏文本、翻译等新功能
- 部落|excel固定显示行列视频:应用冻结窗格同时固定标题行和列