维真生物助力
病毒产品:AAV Ctrl & AAV-MCL-1
注射动物:6月龄雄性转基因APP/PS1 (C57BL/6)小鼠
病毒用量:1.5 μL (0.15 μL/min)
注射方式:立体定位注射
注射部位:海马区
检测时间:30天后
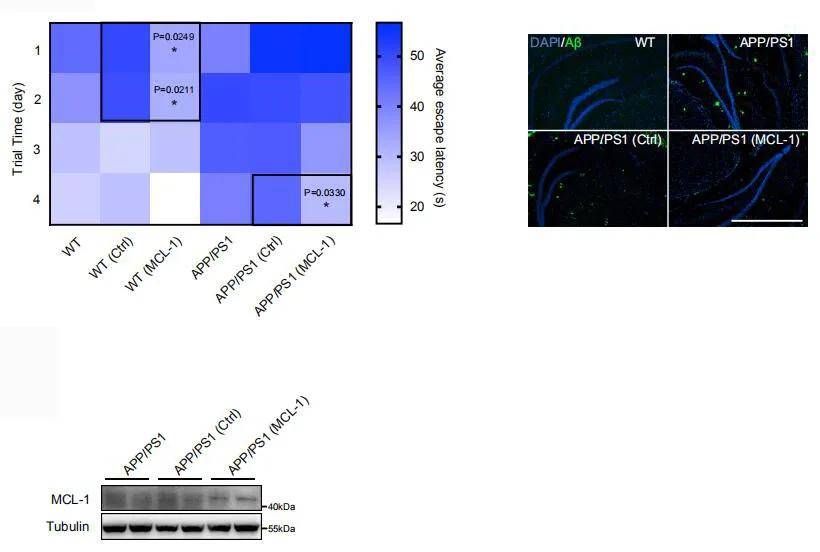
作者以阿尔兹海默症转基因小鼠模型APP/PS1为研究对象 , 将AAV-MCL-1过表达载体注射至小鼠的海马区 , 结果显示 , 过表达MCL-1基因能显著改善APP/PS1小鼠的认知功能 , 而且海马区细胞外的Aβ斑块出现减少 。 此外 , MCL-1的过表达也提高了野生型小鼠的学习和记忆能力 , 这表明MCL-1能在神经元中发挥非常重要的作用 。
文章图片
研究背景
阿尔茨海默病(Alzheimer’s disease, AD)是一种由神经元死亡而造成的神经变性疾病——以进行性记忆力减退和获得性知识丧失 , 直至日常生活活动能力完全丧失为特征 , 给社会和家庭带来沉重负担 , 成为严重的社会和医疗卫生问题 。 阿尔茨海默病是继心血管病、脑血管病和肿瘤之后 , 威胁老年人健康的重要疾病 。 目前 , 全球约有5000万人罹患阿尔兹海默症 。 预计到2050年 , 这个数字将增加至1.52 亿 。 当前 , 全球每年用于治疗、护理阿尔兹海默症病人的费用已经达到1万亿美元 , 而这一数字将在2030年达到目前的两倍 。
阿尔茨海默病的病因复杂 , 目前主流观点认为是由β淀粉样蛋白(Aβ)和微管相关蛋白Tau沉积造成神经元大量死亡引发的 , 1998年以来 , 有100余种治疗此病的药物进行临床试验 , 但仅有6种针对此病症的药物获得FDA的批准上市 , 而且近年来世界各大制药公司针对Aβ或Tau蛋白开发的药物均遭到了不同程度的失败 , 这给人类对于AD的攻克埋上了一层巨大的阴影 。
线粒体功能障碍是AD的一个基本病理特征 , 在散发性和家族性AD病例以及AD动物模型中 , 均被发现有受损的神经元线粒体的积累 。 功能受损的线粒体会触发能量应激 , 从而促进Aβ寡聚化和Tau过度磷酸化 。 线粒体合成调节因子PGC-1α的表达变化以及线粒体功能失调导致的钙稳态失衡都已被证实与AD的发生有关 。 新的研究表明 , AD患者的脑细胞中线粒体自噬受到损害 , 能造成大量损伤性线粒体积累 , 导致突触功能障碍和认知功能的下降 。 因此 , 保障AD患者神经细胞中线粒体自噬的正常进行至关重要 , 而找到一种诱导线粒体自噬的药物靶点则是重中之重 。
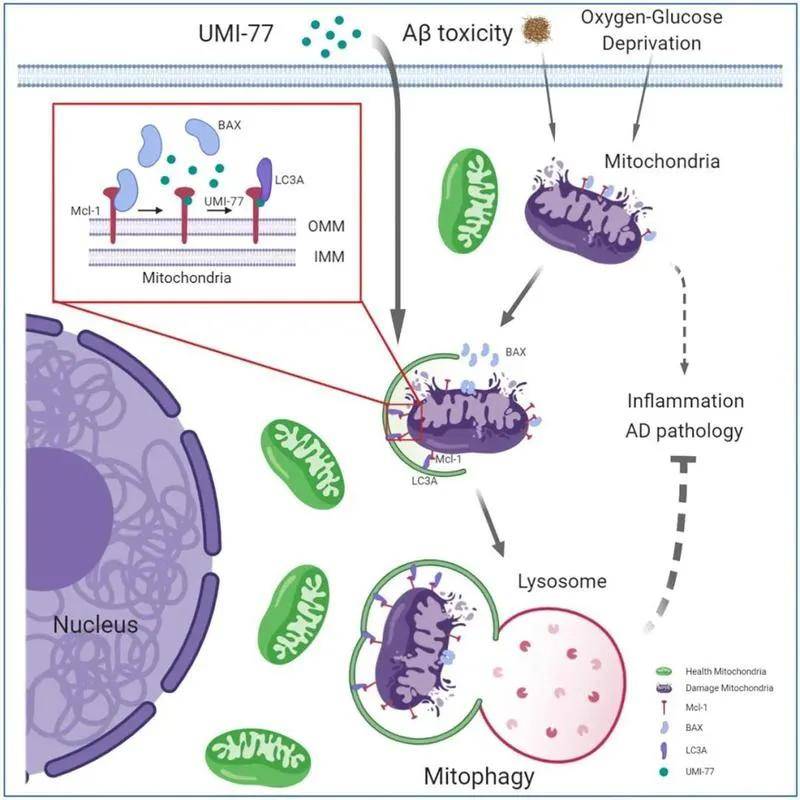
近期 , 浙江大学基础医学院夏宏光教授团队在《Nature communications 》(IF=12.121)上在线发表了一篇关于阿尔茨海默病的最新研究成果—Pharmacological targeting of MCL-1 promotes mitophagy and improves disease pathologies in an Alzheimer’s disease mouse model 。 该研究首次揭示了抗凋亡蛋白MCL-1作为线粒体自噬受体蛋白介导线粒体自噬的新机制 , MCL-1的特异性抑制剂UMI-77可以在AD模型小鼠中显著缓解阿尔茨海默病的病理特征 , 改善小鼠认知;本研究提出靶向MCL-1蛋白诱导线粒体自噬是一种有巨大前景的治疗阿尔茨海默症的策略 。
文章图片
【蛋白|更进一步!!解密阿尔茨海默病治疗新靶点!】 UMI-77作用机制示意图
研究结果
首先 , 研究人员通过利用荧光蛋白Keima构建敏感的线粒体自噬定量检测方法 , 对稳定表达mt-Keima(Keima蛋白具有在酸性和中性pH中荧光信号不同的特性 , 定位于线粒体中的Keima(mt-Keima)可显示通过自噬途径进入溶酶体中的线粒体 , 直观地反映线粒体自噬程度)的HEK293T细胞系和一个包含2024个FDA批准的药物或候选药物库进行小分子化合物高通量筛选 , 找到了一种可以安全有效诱导线粒体自噬的小分子化合物——UMI-77 , 一种Bcl-2家族抑制剂 。
研究表明 , UMI-77是抗凋亡蛋白MCL-1的特异性抑制剂 , 能阻断MCL-1和Bax/Bak之间的相互作用 , 从而允许Bax/Bak诱导细胞凋亡 。 研究者通过测定线粒体的去极化及自噬水平 , 发现与CCCP(氧化磷酸化抑制剂 , 影响线粒体的蛋白合成)相比 , UMI-77在亚致死剂量下不会诱导HEK293T和HeLa细胞的线粒体损伤;在HEK293T-mt-Keima中 , UMI-77+CCCP联合使用明显增强了线粒体的自噬水平 , 表明UMI-77不会造成线粒体损伤 , 且能够促进线粒体自噬 , 且能增强CCCP诱导的线粒体损伤引发的线粒体自噬 。 HEK293T-mt-Keima细胞的活细胞成像实验、透射电子显微镜(TEM)观察及western bloting 实验也证实了UMI-77的促线粒体自噬作用 。
为排除凋亡诱导剂对线粒体自噬可能的影响 , 研究者又做了评估实验 。 结果发现 , 药物库中大多数诱导细胞凋亡的药物及pan-caspase抑制剂Z-VAD-fmk(可阻断细胞凋亡)不会影响UMI-77的促线粒体自噬作用 , 这表明UMI-77诱导的线粒体自噬 , 不依赖于凋亡诱导 , 且UMI-77在亚致死剂量下不会诱导细胞凋亡 。 与此同时 , UMI-77处理后 , 巨自噬标记物p62和其他细胞器标记物的表达水平并没有下降 , 这表明UMI-77可特异性诱导线粒体自噬 , 不影响非选择性自噬 。
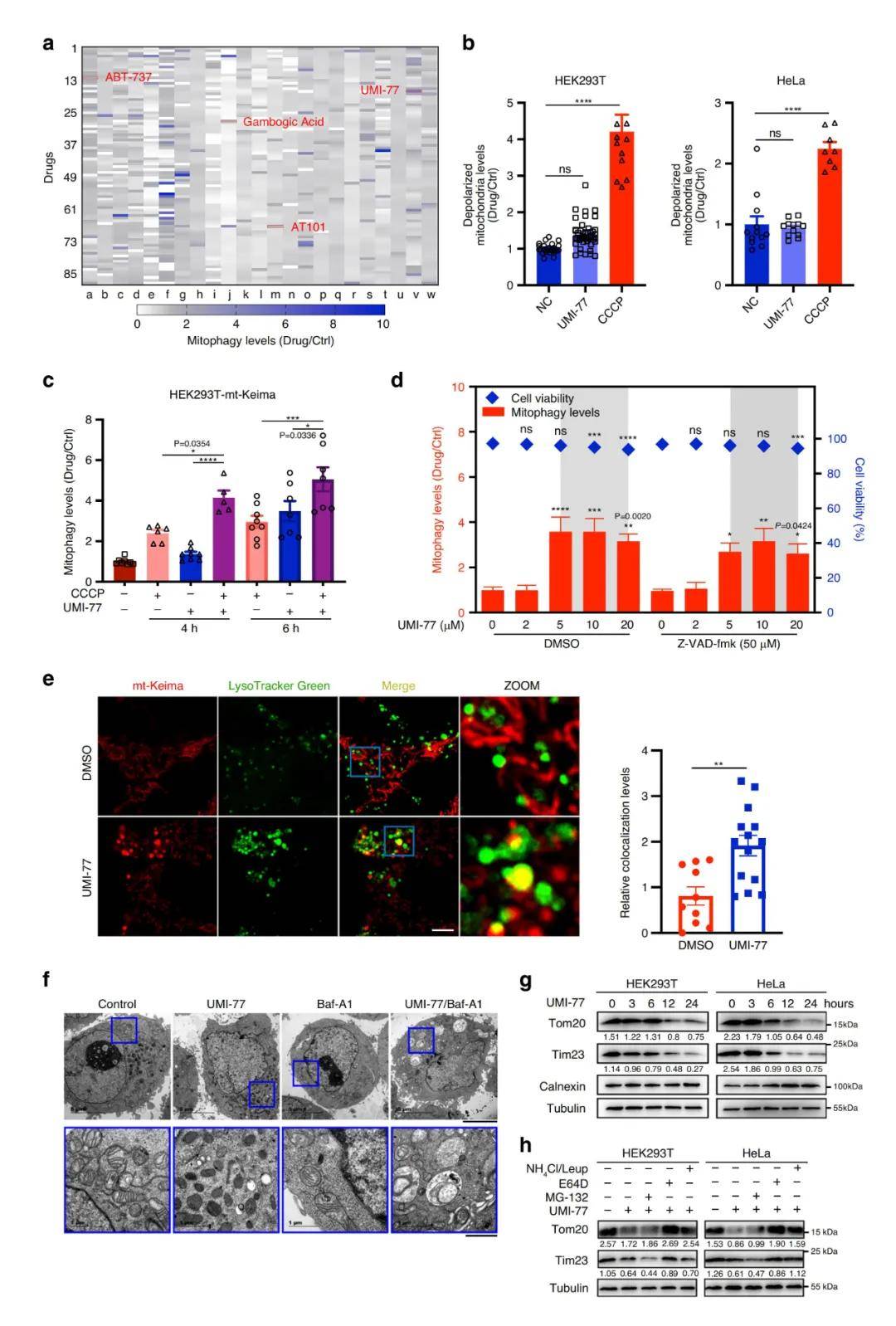
文章图片
图1. UMI-77可特异性诱导线粒体自噬
既然UMI-77能诱导线粒体自噬 , MCL-1又是UMI-77的靶点 , 那么MCL-1是否参与了UMI-77诱导的线粒体自噬的过程呢?研究者对此进行了探索 , 发现在HEK293T和HeLa细胞中敲除MCL-1基因后减缓了UMI-77诱导的线粒体蛋白Tom20和Tim23的降解 , 抑制了UMI-77诱导的线粒体自噬水平 , 这说明MCL-1在UMI-77诱导的线粒体自噬激活中是必须的 。
此前已有研究证明 , 细胞自噬受体如FUNDC1和FKBP8的过表达能促进细胞自噬 , 那么MCL-1在线粒体自噬中是否也是扮演自噬受体的角色?为了探究其在自噬中所起的作用 , 研究者建立强力霉素Dox诱导的MCL-1过表达的HEK293T稳定细胞系(HEK293T- MF2) , 值得注意的是 , HEK293T- MF2中线粒体标志物Tim23的表达水平下降 , 线粒体变小、片段化 , 而且线粒体和溶酶体出现了共定位 。 透射电镜分析也证实了MCL-1的过表达促进了线粒体自噬 。 综上结果表明 , MCL-1在线粒体自噬中发挥关键作用 。
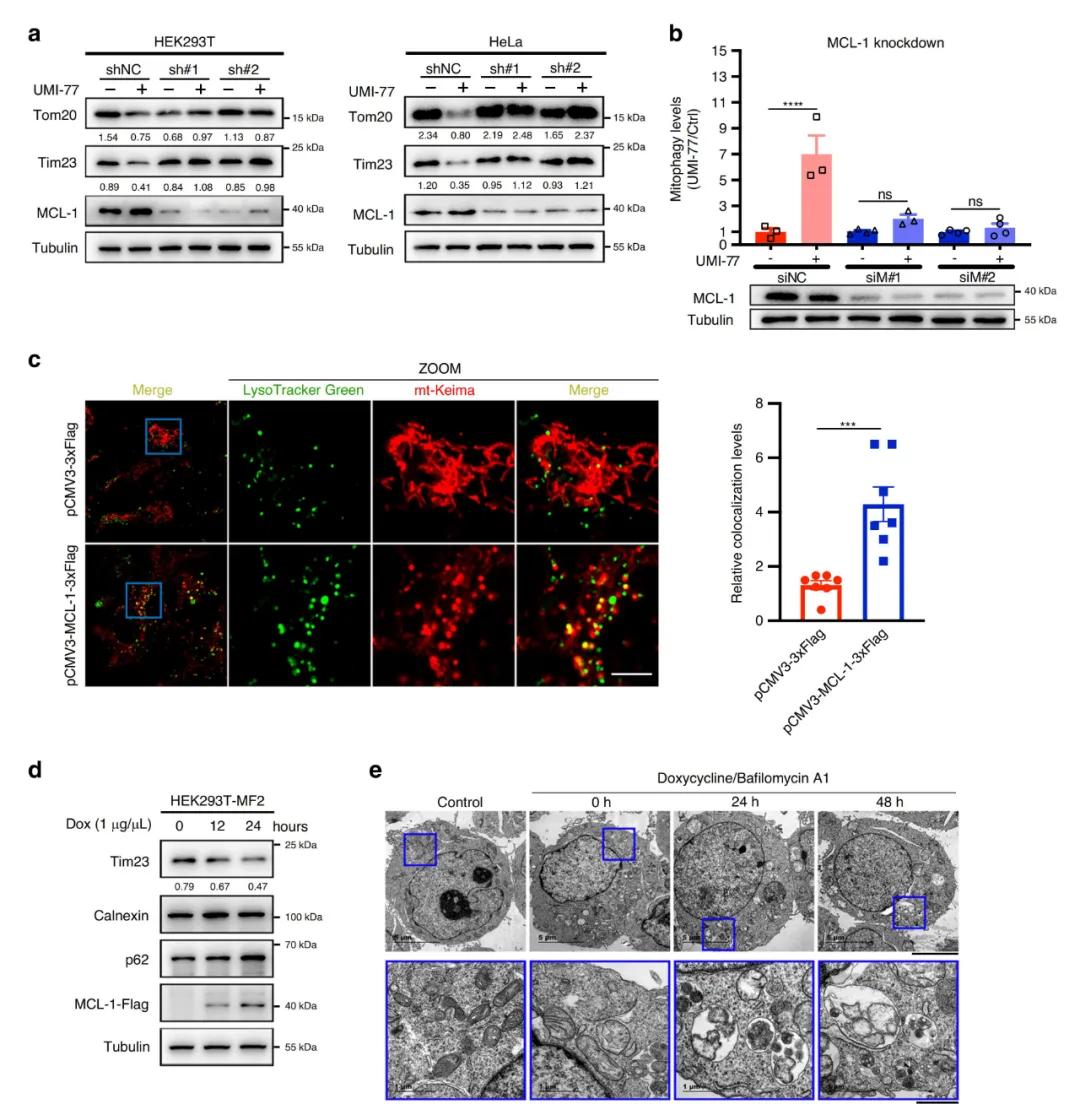
文章图片
图2. MCL-1促进线粒体自噬
研究者对MCL-1在线粒体自噬中是否扮演自噬受体的角色继续展开研究 , 并推测UMI-77诱导Bax/Bak中MCL-1释放后能MCL-1能与LC3发生相互作用 , 从而促进线粒体自噬 。 研究发现MCL-1在其C端有三个“LC3-相互作用区(LIR)”基序 , 其中前两个基序LIR261-264和LIR318-321位于胞质区 , 并且LIR261-264具有很强的保守性 , 这提示它是有一定功能的 。
研究人员首先验证了UMI-77是否能诱导MCL-1和LC3发生相互作用 , 通过免疫共沉淀试验发现 , 经UMI-77处理后 , LC3A与MCL-1的相互作用增强 , 而Bax与MCL-1的相互作用随之减弱;在UMI-77的作用下 , MCL-1还与其他Atg8家族蛋白相互作用 。 此外 , pull-down实验也显示MCL-1能直接与LC3A结合 。 随后 , 研究者又生成了一系列MCL-1 LIR261-264和LIR318 - 321基序的突变体 , 结果显示MCL-1 LIR261-264基序突变后减弱了MCL-1与LC3A的相互作用 。
接下来 , 研究者对UMI-77是否能增强内源性MCL-1和LC3A的相互作用展开了相关研究:结果与我们预期一致 , 发现用UMI-77处理后 , Duolink?PLA结果显示 , 在线粒体上能观察到内源性MCL-1和LC3A相互作用的增强 , 而未用UMI-77处理时这种相互作用也能观察到 , 反映了基础内源自噬的水平 。 这个结果进一步表明MCL-1是一个重要的线粒体自噬受体 。
最后 , 作者又验证了MCL-1和LC3A的相互作用是否能在UMI-77介导的线粒体自噬激活中起作用 。 结果显示 , 敲除MCL-1后 , HEK293T-mt-Keima和SH-SY5Y细胞中UMI-77介导的线粒体自噬水平降低 , 而补充MCL-1/LIR318-321突变体MCL-1的表达可逆转此现象 。
综上所述 , MCL-1是一个线粒体自噬受体 , 它通过其LIR261-264基序与LC3A直接作用 , 并且这种相互作用能被UMI-77增强进而导致线粒体自噬水平的增强 。 此外 , MCL-1和LC3A之间的相互作用是UMI-77介导线粒体自噬激活的关键 。
图3. MIC-1和LC3A的相互作用是UMI -77诱导线粒体自噬所必需的
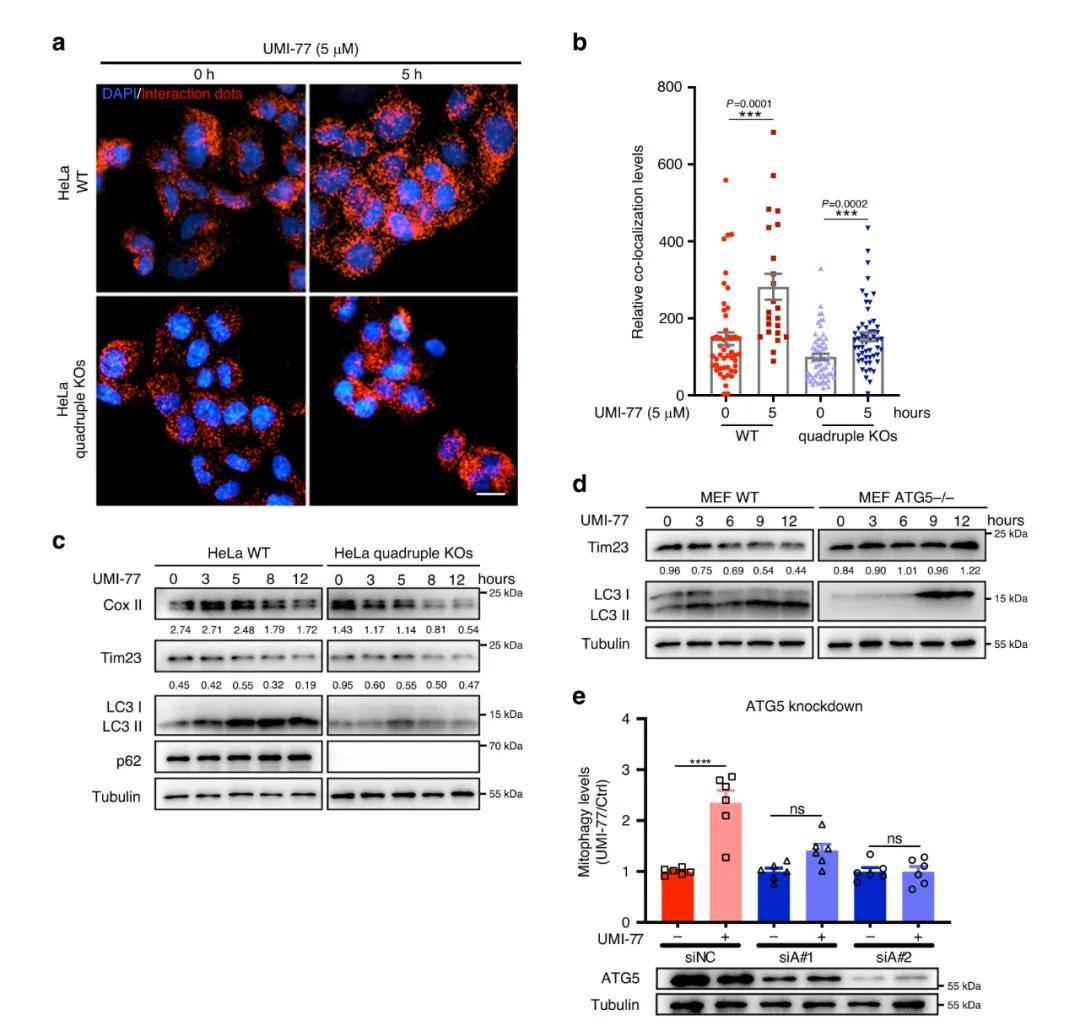
随后 , 作者研究了其他线粒体自噬受体蛋白是否在UMI-77诱导的线粒体自噬或MCL-1-LC3A相互作用的诱导中发挥作用 。 作者以野生型HeLa细胞和四基因敲除的突变体HeLa细胞(敲除线粒体自噬受体NDP52、p62、NBR1和TAX1BP1)为研究对象进行研究 , 发现UMI-77能显著增强MCL-1和LC3A之间的相互作用 , 线粒体标记蛋白Cox II和Tim23的表达水平也均呈现时间依赖性的降低 , 此外 , 线粒体自噬受体(FUNDC1, BNIP3和NIX)也被证明不参与UMI-77诱导的线粒体自噬 , 表明UMI-77诱导的线粒体自噬与这些线粒体自噬受体蛋白无关 。
鉴于UMI-77阻断了MCL-1与Bax的相互作用 , 研究者随后分析了Bax在UMI-77诱导的线粒体自噬中的作用 , 令人惊讶的是 , 敲低Bax后UMI-77诱导的线粒体自噬水平增强 , 说明Bax不参与UMI-77诱导的线粒体自噬 。 另外两种MCL-1相互作用蛋白Beclin1和Parkin也先后被排除参与UMI-77介导线粒体自噬激活的可能性 。 相反 , 在HEK293T-mt-Keima细胞或MEF细胞中敲除ATG5后抑制了Tom20和Tim23降解 , 说明ATG5是UMI-77诱导线粒体自噬必需的 。
综上结果表明 , UMI-77诱导的线粒体自噬是通过ATG5自噬通路介导的 , 不依赖于线粒体自噬受体蛋白NBR1、TAX1BP1、p62、NDP52、FUNDC1、BNIP3、NIX以及MCL-1相互作用蛋白Bax、Beclin1和Parkin 。
文章图片
图4. UMI-77以ATG5依赖的方式诱导线粒体自噬
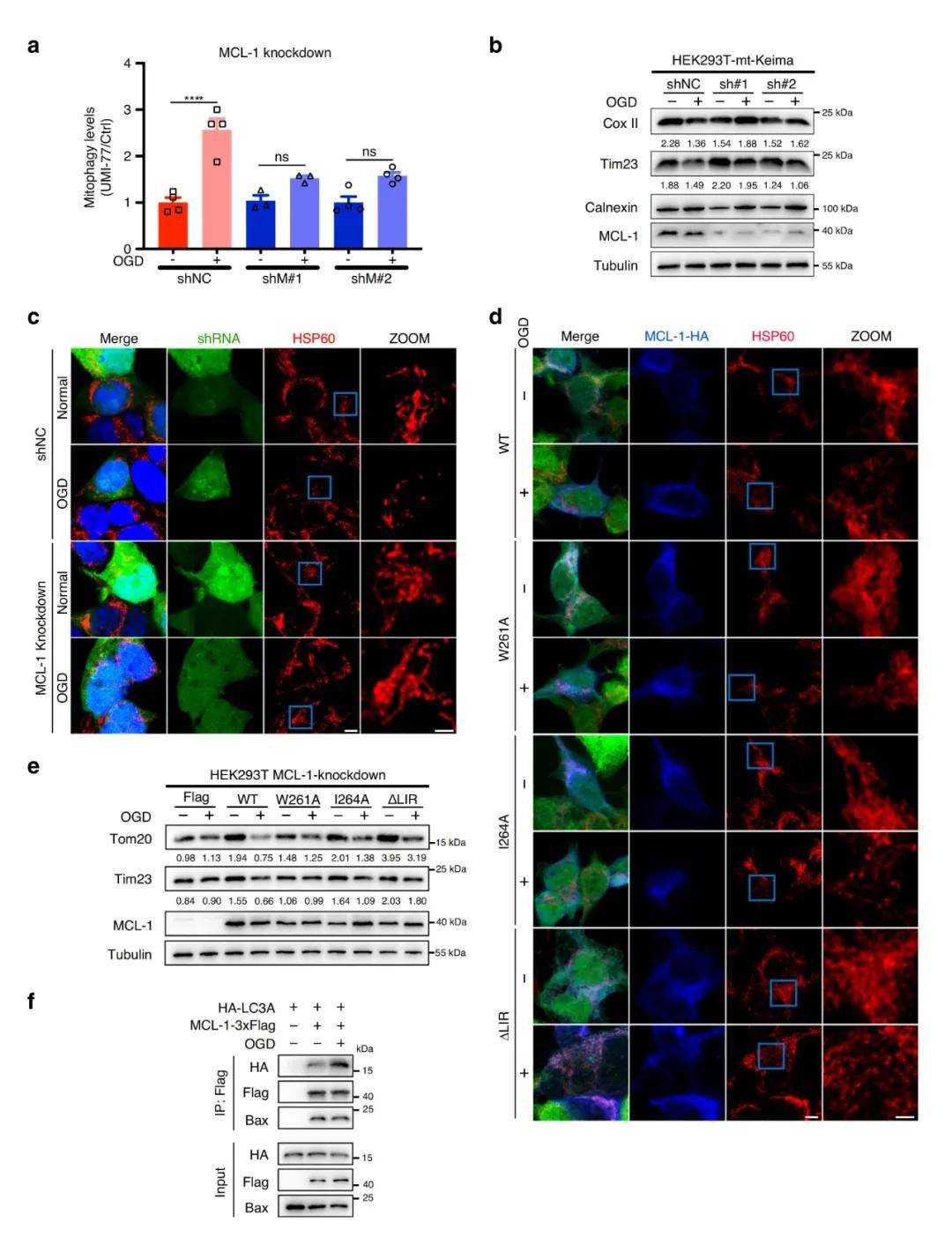
接着 , 研究者又研究了MCL-1作为线粒体自噬受体的生理功能 。 研究表明 , OGD(氧-糖剥夺)或OGD/再灌注会损伤线粒体 , 并通过线粒体自噬诱导清除受损线粒体 。 mt-Keima实验结果显示 , 敲除MCL-1后OGD诱导的线粒体自噬水平显著降低 , 线粒体标记蛋白Cox II和Tim23的特异性降解也被抑制 , 说明MCL-1对OGD诱导的线粒体自噬是必需的 。
已有研究证明MCL-1可调节线粒体的碎片化 , 因此研究人员试图了解MCL-1在OGD诱导的线粒体自噬中的作用 。 免疫荧光显微镜观察到OGD处理后HEK293T细胞线粒体碎裂 , 而敲除MCL-1改变了这些形态学变化 , 表明MCL-1在OGD诱导的线粒体碎片化中起着关键作用 , 并且MCL-1 LIR261-264基序仅参与线粒体自噬 , 而不参与MCL-1的线粒体碎片化调节作用 。 同样地 , OGD也增强了MCL-1与LC3A的相互作用 , 降低了其与Bax的相互作用 , 再次证明MCL-1是一种线粒体自噬激活的受体 。
文章图片
图5. MCL-1是OGD诱导的线粒体自噬所必需的
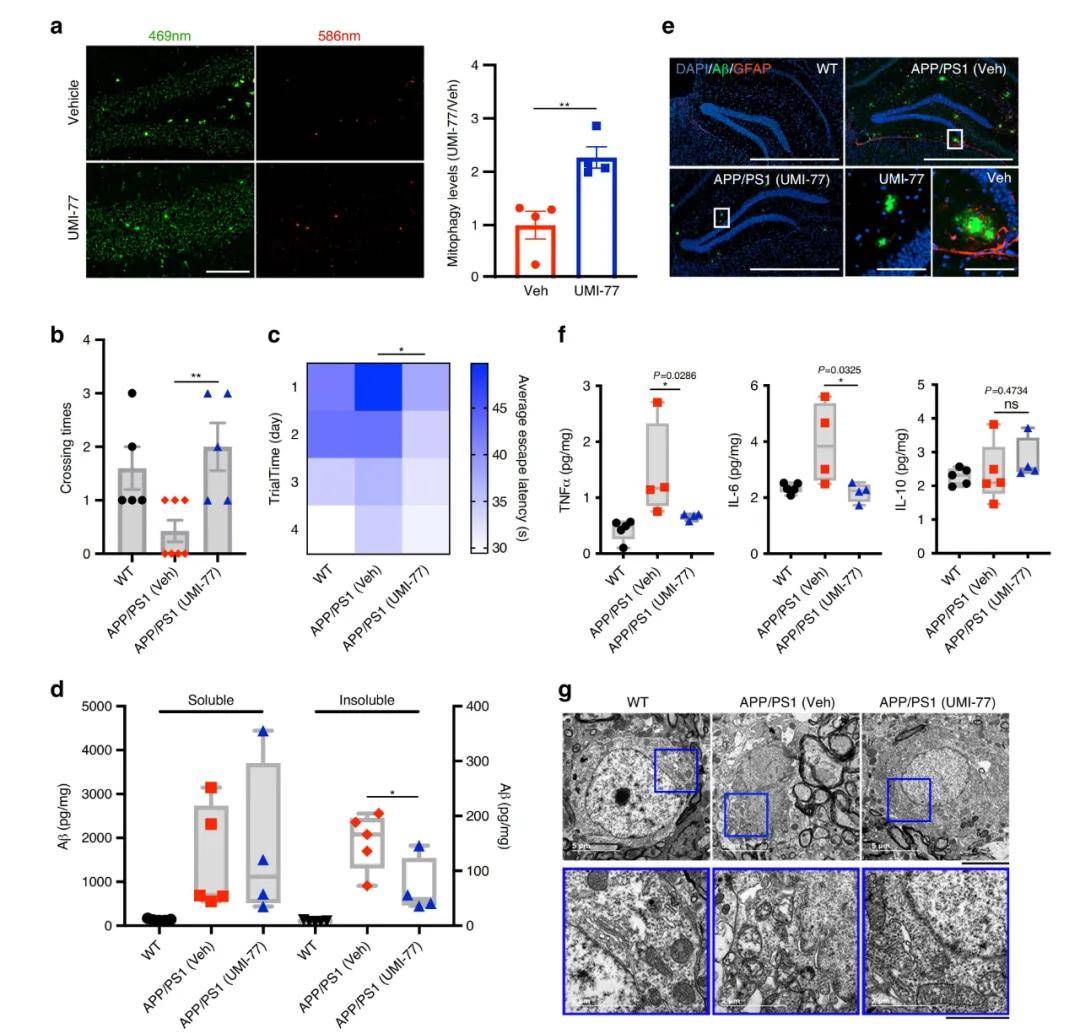
接着 , 研究人员探究了UMI-77诱导的线粒体自噬对AD模型APP/PS1小鼠疾病病理和行为表型的影响 。 对4月龄的小鼠腹腔注射10 mg/kg剂量的UMI-77 , 共持续4个月 。 通过Morris水迷宫测试发现UMI-77治疗明显改善了APP/PS1小鼠的学习和记忆能力 , 且能有效降低小鼠脑内不溶性Aβ1-42的水平 。 同样 , 免疫荧光结果也显示UMI-77治疗后海马区细胞外Aβ斑块明显缩小 , 星形胶质细胞的激活也受到抑制 。 此外 , UMI-77也降低了APP/PS1小鼠的神经炎症水平 , 表现为炎症细胞因子(TNFα和IL-6)水平显著降低 。 重要的是 , UMI-77显著恢复了神经元的线粒体形态 , 这一研究结果与在APP/PS1小鼠中观察到的通过UMI-77可以诱导线粒体自噬从而清楚受损线粒体相一致 。 这些结果均表明UMI-77是一种治疗AD的有效药物 。
文章图片
最后 , 研究人员评估了MCL-1介导的线粒体自噬对APP/PS1小鼠行为表型的影响 。 将AAV-MCL-1过表达载体注射小鼠的海马区 , MCL-1的过表达显著改善了APP/PS1小鼠的认知功能 , 并减少了海马区细胞外的Aβ斑块 。 令人惊讶的是 , MCL-1的过表达也提高了野生型小鼠的学习和记忆能力 , 表明MCL-1在神经元中有非常重要的作用 。
综上所述 , MCL-1是一种新的线粒体自噬受体蛋白 , 是治疗阿尔茨海默症的新药物靶点;UMI-77可以通过释放游离的MCL-1蛋白诱导线粒体自噬 , 显著恢复APP/PS1 AD小鼠模型的认知功能缺损 , 减轻炎症反应和Aβ斑块引起的病理效应 , 促进受损线粒体的清除 。 总而言之 , 诱导线粒体自噬是治疗阿尔茨海默症的有效策略 。
文章图片
推荐阅读






- 蛋白|二代新冠疫苗来了!打了一代,还需要它吗?
- 大豆|烟台双塔食品股份有限公司 低钠高钙零大豆过敏原豌豆分离蛋白研究及产业化通过科技成果鉴定的公告
- IT|阿联酋卫生和预防部批准紧急使用国药第二代重组蛋白新冠疫苗
- 蛋白|血浆蛋白+氧化石墨烯能打印人造肌肉
- 人工智能|人工智能预测蛋白质结构
- 蛋白|中国科学家成功解析植物中独特的双链RNA合成机制
- 实验|AI预测蛋白质结构,正在加快新药研发进程
- 厦门|现场人气爆棚 Cocos 厦门沙龙一站式解密游戏开发关键
- 人员,管理,接触者,人员|一图了解密接人员如何管理、如何解除!
- 标题|一图了解密接人员如何管理、如何解除!